The University of Exeter team visited Ireland this week as part of their ongoing investigation into the biology of the Brent Goose. This species has a remarkable migration, spanning from Northern Canada to Western Europe. The team collects DNA samples, blood for stable isotope analysis and various morphometric and behavioural data. We joined them on Wednesday to help out.
Step 1. Man the netStep 2. Recover your gooseStep 3. Measure your gooseStep 4. Admire your goose Step 5. Release the geese
In my on-going attempt to improve myself I recently attended a lecture by Paul Eric Aspholm on brown bear tracking. This was a much more enjoyable and informative lecture than the last one attended and full of such interesting facts that I just had to share them!
Though the brown bear (Ursos arctos) is found throughout Eurasia and North America the talk focussed on those living in Scandinavia (Norway, Sweden, Finland) and Russia west of the Ural Mountains. In the first half of the 19th century Scandinavia implemented a cull with the intention of wiping bears out. This cull was halted in the 1930s and since then populations have rebounded. Much of the work by Mr Aspholm involved tracking bears to determine population size and movements across the region.
The focus of the talk was shit. Lots and lots of shit. Spring shit, which is mostly grass. Summer shit, which is mostly meat and bones (growing bears need calcium!) and autumn shit, which is mostly berries. The bears are so dextrous that they can pick and eat individual berries and can get through tens of kilos of blueberries in a day! Female bears are quite careful about where they defecate but the males are much more casual in their toilet habits, to the extent that they sometimes accidently walk in a previous bear’s deposit!
The shit is gathered by helpful hunters (who sometimes collect human shit by accident!) and is sequenced to obtain genetic information about the depositor which can be used to track individual bears. Other tracking methods involve collecting hair in hair traps (a line of barbed wire set in a square with a delicious – to bears – smelling lure in the middle) and collecting footprints in snow. This last method was the most innovative (to me, at least). Bears walking over snow will leave skin cells in their footprints. All the scientists have to do is collect a few footprints (around 5-7), melt the snow, get the cells and then sequence the DNA to get a complete genetic profile.
These methods enable researchers to track the habits of individual bears over many years and sometimes several countries. They have been able to show that the bear populations have rebounded well since the end of the cull, with around 8 females reproducing each year in Norway. The hope is to get that to around 15 females a year, still significantly lower than the estimated pre-cull population but a healthy size considering the amount of habitat currently available to them. Most surprisingly of all, the populations show no evidence of genetic bottlenecking or reduced even genetic diversity despite little movement between populations.
As a public service we were given some tips in case we ever come across a brown bear. The traditional advice is to sing. This is, surprisingly, true as it is a sound that only humans can really make. If you scream, you sound like a frightened animal and a perfect chance for a snack. If you shout, you sound like an aggressive animal looking for a fight and the bear is likely to oblige. We were advised not look into their eyes, and most importantly of all – don’t turn your back on it. This is apparently an invitation to play and bears play rough!!
It was an absolutely fascinating talk and I learned so much. Many thanks to Mr Aspholm for such an interesting lecture.
Nazinga Game Ranch is a protected area in southern Burkina Faso, dominated by clear shrub and woody savannah and home to one of the most important elephant populations of the Western Africa.
Researchers from the University of Gembloux Agrobiotech in Belgium tested one of the first unmanned aerial surveys to study the wildlife of Nazinga. They achieved this study using ‘drone’ technology i.e. a small Unmanned Aircraft System (UAS) (pictured). This technology was shown to have the potential to be a valuable alternative to current walk and light aircraft survey techniques.
The Belgian researchers tested different aspects of this new technology on the wildlife and more particularly on elephants. Firstly they wanted to know if the animals reacted when the UAS passed over and found no animal flight or warning reactions were recorded when the plane passed over at a height of 100 meters. Secondly they flew the UAS at different heights (from 100 meters to 700 meters) and showed that only elephants are visible at these heights (while the medium and small sized mammals are not). The pictures taken at a height of 100 meters do however allow easy observation of the elephants.
In the light of this information one elephant survey has been completed in Nazinga Game Ranch so far. This UAS aerial survey has revealed several advantages in comparison to the traditional plane based surveying: (1) an easier flight implementation as a very short airfield is needed, (2) low safety risks as there is no pilot on board, (3) higher reliability in rough weather conditions, and (4) a lower global cost. However, to be able to cover hundred kilometres at a time it is important to improve the flight time of the small UAS as for the moment it is quite low.
Technological improvement of some aspects of the drone will make it more efficient and in the future could compete the light aircraft to monitor the wildlife in Africa.
Author
Florence Hecq: fhecq[at]tcd.ie
Photo Credit
Vermeulen C, Lejeune P, Lisein J, Sawadogo P, Bouché P (2013) Unmanned Aerial Survey of Elephants. PLoS ONE 8(2): e54700. doi:10.1371/journal.pone.0054700
At the launch of our recent college Green Week, Trinity College presented the final stages of its bid to secure the Green Flag Award. Part of the assessment comprised a summary of the plants and animals which, along with the rarefied species of Drama studientis and the Lesser Spotted Theoretical Physicist, contribute to campus biodiversity. Foxes were included in this list which surprised me since I had never come across one campus.
Happily though, last Friday evening one made an appearance just in time for the end of Green Week. Displaying the characteristic “boldness” of its habituation to city life and unperturbed by the passing cars, bikes, and rugby players, a large, healthy-looking fox trotted along the road beside me and into a small patch of scrubby bushes outside the Physics building. It must be a member of the den that resides in the Provost’s Garden – which received celebrity status in a recent Irish Times article. It’s intriguing to speculate whether the Trinity foxes cavort on the cricket pitch long after the last reveller has left the Pav on a Friday night? Similarly, I would love to know whether they are exclusive Trinity residents or do they dodge the shoppers on Grafton Street to visit their cousins in St. Stephen’s Green? Perhaps they also visit Merrion Square – pausing along the way to pay homage to some long lost relatives entombed inside display cases within the Natural History Museum.
Urban foxes have received some bad press recently after the rather gruesome story of the fox which bit off a baby’s finger in south east London. The RSPCA was quick to stress that, while truly horrific, this incident was extremely unusual. Despite their reputation for pilfering unguarded bins, foxes are usually quite shy and wary of coming too close to humans. However, in the wake of the London attack, Mayor Boris Johnson, labelled urban foxes as a “pest and a menace” and there were many calls for a large-scale culling operation to be instigated.
These emotive responses to an isolated incident should not be allowed to dictate future policy for dealing with urban foxes. In his recent New Scientist article, Stephen Harris points out that we are more likely to be attacked by pet dogs rather than foxes and culling programs simply don’t work since new animals just move in to fill vacated areas. In his view, it’s human rather than fox behaviours which give cause for concern. He argues that natural history programs which show cavalier presenters coming in to close contact with wild animals encourage people to seek unnatural and sometimes dangerous proximity with urban wildlife. For example, leaving food out in the garden to attract foxes can lead to some great sightings of these beautiful mammals but placing that food close to a house or near open windows or doors is just asking for trouble. Moreover, feeding foxes is a divisive issue in itself – is it akin to leaving food out for birds or does it equate to just attracting unwanted pests into our gardens? Personally, I have no issue with leaving out scraps but buying cat or dog food just for foxes seems excessive, especially when our untidy cities are veritable all you can eat buffets for these city slickers.
Whether you regard them as pest or surrogate pet, foe or friend, foxes are an inescapable feature of urban landscapes. With Trinity’s campus as their playground, who knows what the one I saw gets up to after dark?
My post on the problem of consciousness troubled a few readers because I dared toy with the idea of dualism, something so offensive to scientists I’m wary to speak its name. But I’m going to continue to argue for dualism because it’s not clear to me that it is wrong even for all the flack it has received. I think a return to this topic is also warranted because of the controversy generated by Thomas Nagel’s latest book, ‘Mind and Cosmos’.
A charge made against my previous post was that dualism is a pernicious idea. Yet nihilism is a negative, and I would argue, damaging philosophy par excellence, but that has no bearing on its truth or falsity. Similarly, one commentator spoke about dualism being argued for because it speaks to a human desire to be something more than physical. But, again, that does not mean it’s wrong. This holds for any idea. We can only take people to task in arguing for something if the only reason they do so is because it comports with their views.
(On a side note I reckon the implications of a physicalist universe are far more terrifying than a dualist one. They are best spelled out by the atheist philosopher Alexander Rosenberg in his essay ‘The Disenchanted Naturalist’s Guide to Reality’.)
Previously I used David Chalmers’ zombie argument to question if the world as we conceive it can account for the presence of consciousness. Chalmers summarises the message of his original zombie argument as “If any account of physical processes would apply equally well to a zombie world, it is hard to see how such an account can explain the existence of consciousness in our world.” Arguments from logical possibility are contentious but let’s not lose sight of what Chalmers is saying.
The subtitle of Nagel’s book is likely to grab the attention of biologists – ‘Why the Materialist Neo-Darwinian Conception of Nature is Almost Certainly False’. But you should hold your cries of “Creationist!” because they’re misplaced. I really need to stress that this debate should not be framed as being between science and religion or science and pseudoscience but rather physicalism and dualism. Indeed some of the more prominent defenders of the latter position, Nagel and Chalmers included, are card carrying atheists.
I haven’t read Nagel’s book yet (I hope to do so) but you can get a sense of the themes he develops from the following description, “The modern materialist approach to life has conspicuously failed to explain such central mind-related features of our world as consciousness, intentionality, meaning, and value.” He argues that there is something more needed to get us from physical matter to conscious thoughts, not even evolution by natural selection can get us there with a purely material world to manipulate. There is a difference of kind rather than degree here.
Some of the criticisms of his latest work argue that he leaves a lot unsaid and many of his arguments have been criticised as vague. But Nagel is most famously known for a famous 1974 essay he wrote on ‘What Is it Like to Be a Bat?’ He says that although bats are conscious they experience the world in an entirely different way to us i.e. there is something like it is to be a bat. If we imagine ourselves as being bats it’s actually through our human minds, i.e. we’re imagining what it would be like for us to have echolocation, which is entirely different. If instead we record all the sensory data that a bat experiences we’re still left wondering about how he/she experiences the world from its own point of view. The normal physicalist approach leaves us guessing.
Previously we were asked to have a prior commitment to physicalism but there are a number of properties of consciousness that should cause us to at least re-evaluate our priors. Neurons, physical entities that they are, do not seem to have the tools to do what is being asked of them. When it comes to physicalism, past performance is no guarantee of future success.
I would like to say that the talk presented a range of evidence for intelligent design and carefully countered the usual arguments against it. I would like to say this, but I can’t. The talk, which lasted over one hour, spent much of the time quoting non-scientists and misquoting scientists, painting ID proponents as martyrs to the cause and science as tautologically incapable of addressing questions of design. The religious beliefs of ID proponents were constantly referred to, despite supposedly being completely irrelevant, which was an indication that this was, after all, a religious proposition not a scientific one.
It would be easy to question the credentials of Dr Alistair Noble (PhD in chemistry) and ask how someone who has been outside of scientific academia longer than I have been alive can claim to have found fundamental flaws that no working biologist has been able to find, but I won’t. Instead, I have tried to focus on the claims of Dr Noble and see if they can be answered (see my last blog post).
There is much more that I could have said. The case for evolution is so strong that I could go on for hours about the evidence from multiple disciplines that support it. It seems that the same cannot be said for intelligent design. Dr Noble spent about 15 minutes of his (more than) one hour talk providing evidence which can be easily refuted by anyone who has even a basic understanding of evolutionary theory. His ‘evidence’ ultimately boiled down to an Argument from Incredulity with a side helping of the Argument from Authority.
I was disappointed by the lack of scientific rigor Dr Noble exhibited. Not one journal article was presented, not a single claim that hasn’t been refuted multiple times before. I had hoped for an intellectually stimulating talk that would force me to question my understanding of evolutionary theory but instead I was confronted with the same, tired claims that have been presented by ID proponents for years now. It is a shame that Dr Noble could not have used his clearly considerable intellect to study the actual science and see that evolutionary theory is not a threat to his faith but is an amazingly simple yet profound explanation into how the diversity of life arose.
On the 31st January, stimulated by a European Food Safety Authority report, the EU proposed banning three neonicotinoid insecticides which have been implicated in causing honeybee decline. These insecticides are widely-used, systemic (i.e. soluble enough in water to move around the plant’s vascular system to nearly all plant tissues), and, like nicotine, affect the insects’ central nervous system. They are highly effective at reducing insect pests that feed on crops and reduce yields and value, and many farmers are concerned about the effect the proposed ban will have on crop production. But these insecticides can also end up in the nectar and pollen of crops (as well as in the soil and in non-crop plants), and thus can have unintended side-effects on beneficial, nectar-feeding insects, who act as pollinators. Especially bees.
Bee decline has become a hot topic with scientists, the media, the public and even some politicians, but until recently the threat of neonicotinoids to bees has not been seriously implicated in their decline. Concern about pollinator decline is a result of the important role that pollinators play in food production: 75% of crop species depend on animal pollinators, which translates into 35% of global production; and the total annual economic value of pollination has been estimated at €153 billion globally. In addition, pollinators are fundamental to most terrestrial ecosystems, and indirectly affect the availability of food for other organisms (e.g. fruits and berries for frugivorous birds), as well as the structure and functioning of ecosystems.
So here’s the paradox: flower-visiting insects including bees are really important for agricultural production. But so is the use of neonicotinoid pesticides. Which is more important and is the ban justified on scientific grounds?
In the last year, the evidence that neonicotinoids have negative impacts on bees has been mounting. Bees and other flower-visiting insects are exposed to neonicotinoid pesticides in multiple ways: during planting of seeds which have been coated with pesticides as a pre-planting treatment, by collecting pollen and nectar from the crop, and by foraging on non-crop plants which take the pesticide up through the soil. Traditionally, toxicological tests of agrochemicals are carried out on the managed honeybee Apis mellifera, and pesticides are rated according to their lethal effects (by calculating the LD50 – the dose required to kill half the organisms tested after a specified duration). But the biology of Apis and all the other bee species (20,000 of them worldwide) is different. Can we generalise about effects on Apis, to effects on other bee species, and other pollinating insects including hoverflies and butterflies? And what about sub-lethal effects, i.e. those that don’t kill the insects, but affect their physiolology, behaviour and fitness?
Neonicotinoids are highly toxic to insects – that’s the whole point of them. Bees are insects. So it shouldn’t be too much of a shock that they kill bees. Last year it was shown that neonicotinoids can also have sub-lethal effects in honeybees, by decreasing foraging success and navigation by individuals back to the hive. At the same time, the neonicotinoid pesticide, imidacloprid, can reduce bumblebee colony growth and fitness by affecting their feeding behaviour. Some dissenters have cast doubt on the field-relevance of laboratory tests, claiming that field-realistic dosages have not been used, but this is not the case – the concentration of imidacloprid in oilseed rape flowers for example has been found to be 4.4-7.6 mg/kg in pollen and 0.6-0.8 mg/kg in nectar, which was within the range tested on bumblebees. This is pretty convincing evidence that neonicotinoids can cause very adverse effects on populations of these social bees.
Although neonicotinoids are not the only cause of widespread bee decline, they are more than likely contributing to it. Some of the agrochemical companies are claiming that bee decline has nothing to do with their chemicals and instead blame decline on Varroa destructor, the parasitic mite which infects honeybee colonies. Whilst Varroa probably plays its part in honeybee decline, the most probable cause of decline in other bee species is multiple pressures, including habitat loss and loss of forage plants, AND the use of neonicotinoid pesticides.
So should these insecticides be banned? YES, if we want to address pollinator decline. They should not be used for insect-pollinated crops, and wind-pollinated crops that insects forage on (including maize). But what’s the alternative for the farmer? How can crop production be maintained in the absence of these chemicals? Use something worse? If we’ve learned anything since Rachel Carson’s “Silent Spring” published 50 years ago last year, it’s that an alternative will be found, and we can’t be sure that this won’t be worse for the bees and other pollinating insects.
*by parasites here I am referring to all kinds of infectious disease causing agents including bacteria, viruses, fungi, protozoa, helminths and arthropods.
Why do we care about primate parasites?
Many of the most devastating infectious diseases in humans have origins in wildlife. For example, the global AIDS pandemic originated through human contact with wild African primates and influenza viruses circulate among wild bird populations. These are not only historical occurrences. Recently, for example, rodents were identified as the source of a Hantavirus outbreak in Yosemite National Park, USA . As human populations continue to expand into new areas and global changes in temperature and habitat alter the distributions of wild animals, humans around the world will have greater contact with wildlife. Thus, understanding which infectious agents have the potential to spread from animals to humans is crucial for preventing future human disease outbreaks.
Many efforts are being made to collate information on wildlife and human diseases. Much of my research (which I will blog about when I get chance!) uses an amazing database known as the Global Mammal Parasite Database or GMPD for short. Every time a paper is published which contains details of parasites found in either primates, carnivores or ungulates, the information is added to the database. As much data as possible is recorded, including the species infected, the type of parasite, the prevalence of the parasite, and the geographic location of the study. Prof. Charles Nunn and his colleagues have been collecting data for the GMPD since around 2005 and it currently contains around 6000 records for primates alone. This definitely makes it the most comprehensive dataset of primate parasites in existence.
The GMPD sounds amazing…so what’s the problem?
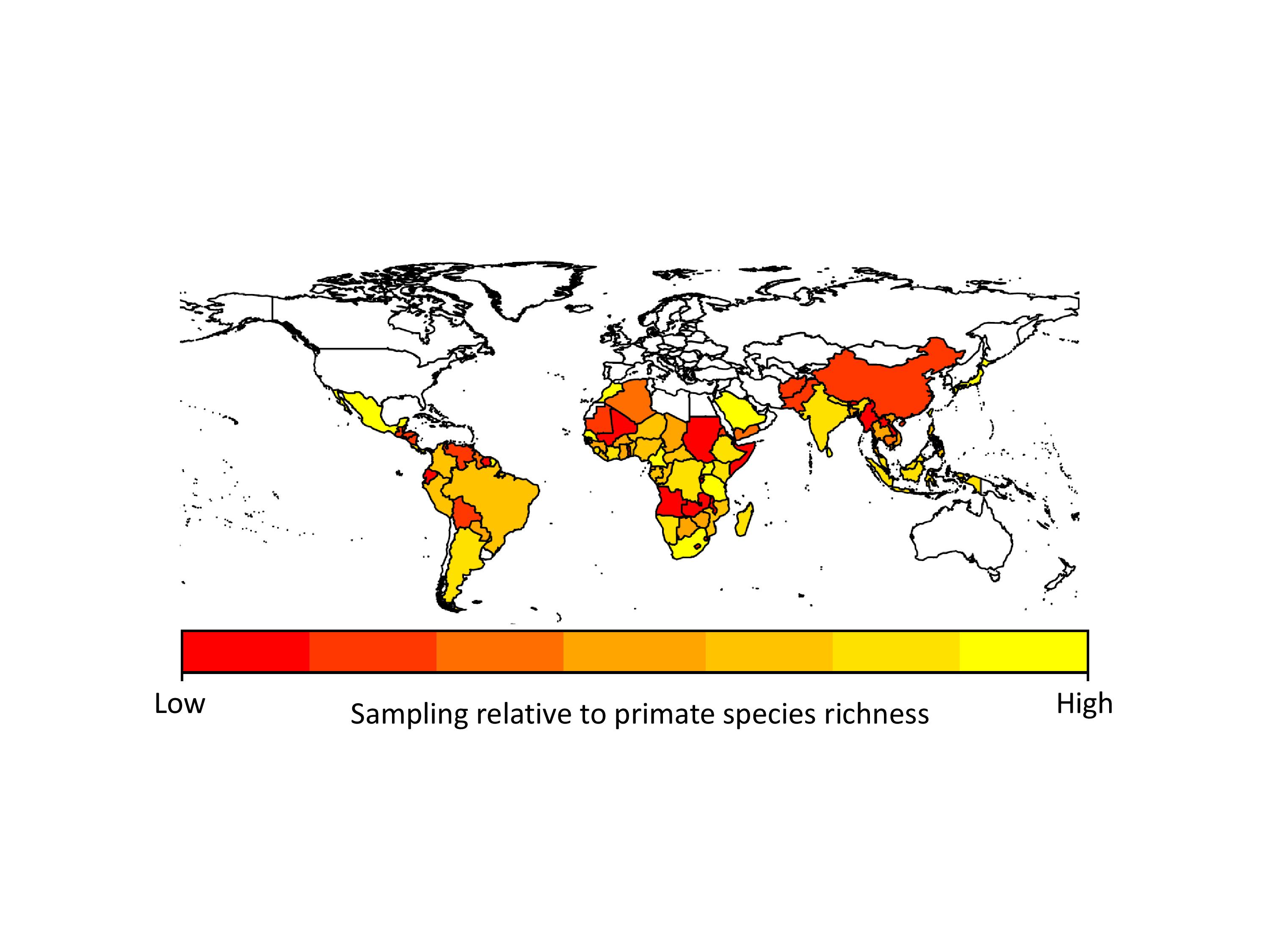
The problem with the GMPD (and this is a feature of virtually all datasets) is that there is sampling bias. Certain primates are sampled for parasites much more frequently than others. Chimpanzees, for example, are sampled for parasites all the time, whereas species such as tarsiers are sampled much less often. This has the effect of making it look like chimpanzees have far more parasites than tarsiers, simply because they have been sampled more often. In analyses using the database we usually deal with this problem by adding sampling effort into our models, so we give less emphasis to high numbers of parasites in primates we have lots of samples for. Unfortunately this problem is also evident when we look at parasites (things like malarial parasites are often sampled because of their importance to human health) and geographic regions (areas with primate research stations are sampled far more regularly than more remote regions). If we hope to use the GMPD data to make reliable predictions about future risks to humans, we need to identify gaps in our knowledge of primate parasites.
So what did you do?
Without going into the technical details, we looked across the primate phylogeny and primate geographic ranges to identify gaps in our knowledge, and used statistical models to investigate what factors led to primates and geographic areas being relatively well- or relatively poorly-sampled. We also used species accumulation curves to extrapolate parasite species richness for primates.
Where are the gaps in our knowledge?
We found that apes (chimpanzees, gorillas and orangutans) were generally better-sampled than other primates, but there was incredible variation in sampling among all other major primate groups. Apart from apes, the primates that researchers appear to sample most are the species they encounter most often, i.e., widespread, terrestrial, diurnal species. However, some primates were sampled more often because they are already intensively studied for other research, because they live in frequently visited field sites, or because of their importance in medical research. Across countries, we found that in general, parasite sampling is highest in countries with more primates to sample. We expected that the GDP of the countries would also affect sampling effort, with wealthier countries having more money for disease monitoring. However, we found no evidence for this in our analyses, probably because most research on primate diseases is not funded by the country in which the research takes place.
Sampling effort for primate parasites across the world. Poorly-sampled countries are in red, and better sampled countries are in yellow.
When we extrapolated parasite species richness values we found that even within our best-sampled primates and countries, we are missing a lot of parasites. On average we predicted that 38-79% more parasite species than currently reported in the GMPD should be found in our best sampled primate species, and 29-40% more parasite species than currently reported in the GMPD should be found in our best sampled countries. This emphasizes exactly how poor our sampling is across all primates and countries. Another concern is that although viruses make up only 12% of the parasites in our dataset, viruses arguably present the greatest zoonotic disease threat to humans because their fast rates of evolution should allow them to easily adapt to new hosts.
What next?
Identifying parasite sampling gaps across primate species and geographic regions is only the first step; we need to find strategies to minimize these sampling gaps if we are to predict which primate diseases may emerge in humans. One solution is to set research priorities based on the sampling gaps, for example, by focusing effort and funding on relatively poorly-sampled primate species, arboreal primates, those with small geographic ranges, or those found in relatively poorly-sampled regions of South East Asia, Central and Western Africa, and South America.
Focusing on relatively poorly-sampled primate species and areas may improve our general understanding of primate parasites, but it is only one factor in predicting risk to humans. For example, hosts are more likely to share parasites with their close relatives than with more distant relatives. Thus, continuing to focus our sampling efforts on parasites of our closest relatives (chimpanzees, gorillas and orangutans) may provide the greatest return in the case of risks to humans. This is particularly important because we found that chimpanzees are expected to have 33-50% more parasites than currently found in the GMPD. In addition, ecological similarities also influence parasite sharing among primates, and humans share more parasites with terrestrial than arboreal primate species. As with sampling effort, this probably reflects higher contact rates among humans and terrestrial primates compared to arboreal primates. As a related issue, a host living at higher density is expected to have higher prevalence of parasites and may have more contact with human populations or our domesticated animals, thus increasing opportunities for host shifts to humans. The large numbers of zoonotic emerging infectious diseases with rodent or domesticated animal sources also highlight the importance of rates of contact and host density for disease emergence in humans.
In conclusion Sampling effort for primate parasites is uneven and low. The sobering message is that we know little about even the best studied primates, and even less regarding the spatial and temporal distribution of parasitism within species. Much more sampling is needed if we hope to predict or prevent future emerging infectious diseases outbreaks.
Recently Science published O’Leary et al.’s – new load of oil to fuel the burning debate on the origins of placental mammals.
Just to be clear: there is an important distinction between mammals in general that includes many fossils from the Jurassic as well as the extant platypus, kangaroo and your grandma; and placental mammals that includes your grandma and the armadillo (but not kangaroos or the platypus) and no fossil before 65Myr. For readers that are not used to the debate concerning the first placental mammal here’s the main question: did the first placental mammals diversify before or after 65Myr (the important KT boundary)?
(1) After Katie
One view suggests that early mammals lived in the shadow of dinosaurs and the demise of these mighty creatures allowed our rat-like ancestors to take over the earth, the seas and the skies “Save yourself mammals”. This idea was proposed by Simpson in the 1950’s and is supported by the fossil record; many dinosaurs (both big and small) were present before the 65 Myr KT boundary, then a catastrophic meteorite impact marked the KT limit and placental mammals radiated after that. This sequence of events seems to be very straightforward but reality appears not to be so simple. Increasing numbers of mammal species from the late Cretaceous are being discovered, (including rather big ones feeding on dinosaurs) and not many species in general are found in the fossil record before 55Myr when all groups of placental mammals seem to suddenly appear (for a full story see Luo’s 2007 Nature review).
(2) Before Katie
In contrast, another group of people, mainly assisted by molecular dating methods, found out that post-KT placental mammalian diversification may just be an artefact of the fossil record (like Meredith et al 2011 in Science again). Their DNA evidence seems to say that placental mammals evolved before the KT limit and that either palaeontologists failed to find them or else the fossil record failed to preserve them. One major criticism that moderate people argue is that there are still problems associated with molecular dating methods. I won’t go into the details (yes I’m trying hard not to) but molecular dating relies on DNA on the one hand (sampling quality and modelling) and on the fossil record on the other hand. So if the people using DNA criticise the fossil record and want to improve the DNA dating estimates, they have to rely on the same fossil record that they are criticising. The snake bites his own tail.
So what about O’Leary and colleague’s paper? They basically support the first theory (placental mammals evolved after KT). Fair enough, it was led by a number of great palaeontologists and based on a massive morphological data set (~4500 characters introduced as phenomics (from the phenotype) as opposed to genomics (from the genome) data) collected on 40 unambiguous fossils and 46 extant placental mammals. Genomic data based on 26 genes of these extant placental mammals was also included. This paper is the result of an impressive and unique collaborative work, but – Ned Stark from Games of Thrones said “nothing someone says before the word “but” really counts” – but this paper is criticisable…
First of all, the data set: although the morphological data is impressive, the taxa sampling effort seems a bit weak, especially for extant placental mammals. Meredith et al used the same genomic data (26 genes) but based on ~164 mammals to answer the same question. Why couldn’t O’Leary use all of this already published mammal DNA? For the second criticism, I’m just going to quote Yoder’s review published in the same issue “Today, sophisticated theoretical and computational methods are used to estimate and calibrate molecular phylogenetic branch lengths (which represent time). Together with improved methods for integrating fossil and molecular data, dates derived from molecular phylogenies have inched closer to those implied by the fossil record. Is the approach used in the O’Leary et al. study directly comparable to these recent molecular phylogenetic studies? Not really, as it turns out.”
No wonder this paper supports the first theory, it is just a precise and massive analysis of the 40 species of the placental mammals fossil record. Personally, I’m really frustrated by how they managed to publish this paper. Since it’s part of my PhD research, I automatically get excited when I see fossils mixing with extant species so I really hoped this paper would link the two approaches instead of supporting the old fashioned view of evolution (the dinosaurs dying and the mammals taking over). I’d like to think that the history of life is a bit more complex and exciting…
A last comment to justify my title and which will be my main critique to this paper is that O’Leary et al. tried to recreate the “hypothetical placental mammal ancestor”.
As I said, this paper could be seen as a summary of the placental mammal fossil record. So why did they break the first rule that keeps palaeontology away from palaeo-poetry (i.e. going too far with palaeontological hypotheses)? Here they reconstructed a whole creature using their morphological data. What they made was essentially a mean (average) placental mammal (a primitive rat-like creature) – a throw-back to the early stages of palaeontological views of mammalian evolution. What did the ancestor of a duck and a beaver look like? Something in between – a platypus for example? As Olaf Bininda-Edmonds said on Ed-Yong’s Nature post “comparing the two estimates is like comparing “apples and oranges”, they haven’t really done anything to resolve this on-going dispute”.
This paper has also caused controversy on twitter. I’ll just cite two opinions.
Gavin Thomas (@Phalaropus)
“The reconstruction is fun – I’d love to see a picture based on 95% CIs for the ancestral states.”
and Rich Grenyer’s answer (@rich_)
“yes indeed. Something like this” (see our title image).
Many parts of the online science community got excited about this paper, you can see further discussions on Jerry A Coyne’s blog (here and here), on Ed Yong’s one (here and here or there) or else on the twitter feed #placental.
At first glance, many scientific ideas can appear counterintuitive. A press release from a leading Irish wildlife charity in support of the proposed coursing ban prompted me to attempt to balance the discussion of coursing impacts on the Irish hare population. The bill to ban coursing is due to come before the Dáil in the coming months. However, the above press release immediately struck me as biased, and so I felt a discussion of coursing impacts was required before the public were asked to sign any petitions in support of this ban. For those unsure of just what coursing is, it is a popular field sport which consists of a hare being chased by a pair of greyhounds over a short distance. Unlike fox and deer hunting, the aim of coursing is not to kill the hare. It is instead a speed and agility competition between two dogs, where each is awarded points depending on its ability to “turn” the hare from a direct route along the field. Irish hares (Lepus timidus hibernicus Bell 1837) are caught and held in captivity prior to an event during which the hare is coursed within an enclosed park. A running hare is given a 75m head start before the release of two dogs, whose performance is assessed by a judge, and surviving hares escape into an area from which the dogs are excluded. The duration of the pursuit is relatively brief, usually lasting less than a minute, and surviving hares are returned to the wild after the event.
The IWT states something which a few of us may agree with; that Ireland is lagging behind in terms of its attitude to welfare and conservation of native wildlife. However, the idea that a coursing ban would in some way improve this status is highly questionable. Welfare issues need to be taken into account, but these considerations must be viewed in parallel with the beneficial aspects of coursing, such as habitat conservation and the associated protection of both target and non-target species, before any final judgements regarding coursing acceptability can be made. It is perhaps unintuitive, but evidence indicates that coursing has an extremely large positive impact on hare numbers. Mortality of coursed hares stands at just 4.1% since the implementation of dog muzzling in 1993, and research has found coursing to have negligible impacts on hare populations due their large intrinsic rates of increase. People who participate in coursing maximise hare populations in coursing preserves through predator control and set aside to conserve habitat suited to the Irish hare. In fact, it is agricultural intensification (an issue completely ignored in the IWT article) which is more likely to blame for population declines. Habitat management to encourage target species for hunting can protect against the detrimental effects of modern agricultural policy on biodiversity. Irish Coursing Club preserves host a hare density 3 times greater than that supported by the wider countryside. What is more probable is that coursing is actually stemming the tide of anthropogenic destruction of many species our native wildlife (including corncrakes and many other farmland bird species) through habitat conservation aimed at artificially increasing hare populations for coursing. If coursing were to be banned in this country, this practice would be completely abandoned due to waning interest in encouraging hare numbers, and could potentially have serious ramifications for other wildlife which benefit from associated habitat management and predator control. Incentives to promote hare conservation would be required, but it’s questionable whether these would produce the same results as coursing-associated management due to a lack of personal interest for farmers and other landowners who practice coursing. Hare conservation in the absence of coursing, similar to that of other species benefiting from game management, would be a costly endeavour and would be unlikely to be awarded the necessary funding in the Republic of Ireland with the current economic climate.
We have the opportunity to be forward-thinking, innovative and inclusive in the way in which we achieve sustainable conservation of our native wildlife, something which appears all the more important in light of the EU agricultural policy reforms which were leaked in recent days. We can only hope that a review of the research will stop the Dáil bowing to ill-informed political pressure and perhaps, the future of farmland birds and our only endemic mammal, the Irish hare, will be ensured.


























{kind=link}