Antler collection at Harvard’s Museum of Comparative Zoology
It may be an inconvenient truth in these conservation-focused times but we owe a debt of gratitude to the trophy hunters; the army officers and colonial-types who killed animals for sport and prestige. Without their considerable efforts, the vaults of natural history museums would be devoid of the skeletons and skins which form the bases of both exhibitions and many PhD and MSc. theses. Of course, were it not for the over-zealous efforts of hunters perhaps many charismatic animal species wouldn’t be so endangered now but let’s focus on the positives here…
Naturally, if you’re a hunter looking for a prize, bigger is usually better. It’s far more impressive to have a stag or bear’s head mounted on your wall than a hedgehog or shrew – although I would like to hear the embellished stories which might arise from tales of killing your first hedgehog! Similarly, if you are a taxidermist or museum collections manager in many ways it is easier to prepare and preserve large rather than small mammal specimens.
These collection trends are all well and good if you’re interested in the charismatic species. I, however, am studying the little shrewy-type things; hedgehogs, moles, shrews, golden moles and tenrecs – fascinating species but not prized possessions for your trophy case. Even in some of the world’s largest natural history museums it’s difficult to find intact skulls and skeletons of some of these creatures. Combined with the inherent delicate nature of these animals (some tenrecs’ limbs are tiny!) compiling a complete morphometric data set of the groups remains challenging.
Tenrec limbs with a pen for scale; teeny tiny tenrecs!
For the museum collections of these little critters that do exist; I am eternally grateful to the progenitors of carefully handwritten labels accompanying the skulls and skeletons on which my PhD research depends. Adventurers such as Major Forsyth, G.K., Creighton, and C.J., Raxworthy who donated specimens from their tropical voyages deserve special thanks in any research which arises from their collective efforts. However, I am also grateful to the back-yard naturalists, the people who collected and preserved the seemingly ordinary, every-day species of common shrews and hedgehogs which are no less important to ecological and evolutionary research than their exotic counterparts. I benefitted greatly from their collective efforts during my recent trip to Chicago’s Field Museum.
Whether naturalists or trophy hunters, the individuals immortalised by museum specimen labels couldn’t possibly have envisaged all of the diverse future research which would be based on their prized collections. It’s an important reminder that, despite the inherent appeal of flagship species, the lesser-spotted or common-something-or-others are just as deserving of our attention and study. Even if you don’t have the equipment or inclination to start posting skeletal remains of common species to your local natural history museum, there are still plenty of ways of contributing to the study of “ordinary” wildlife. Don’t be dazzled by the allure of large-animal trophy hunting and remember that, when it comes to understanding the natural world, bigger is not always better.
The ESEB conference this August in Lisbon was not only about Drosophila and #superbock. Among the useful discussions and the interesting talks, a definite highlight came from our very own Kilkenny scaling man all about time perception and comparative analysis… Argh no I missed that one – apparently there was even a realistic Tiger Beetle hunting impression! There were at least eight overlapping talks at any one time and, as I had already seen Kevin’s talk, I went to listen to Folmer Bokma’s insightful talk instead.
I felt Bokma’s talk was a good follow-up to Gene Hunt’s excellent talk on gradualism vs. stasis which he demonstrated through his careful study of the Ostracod fossil record, recently published in the excellent August issue of Methods in Ecology and Evolution. By means of a lengthy introduction with citation of more or less unknown people such as H. Falconer and C. Darwin, Bokma emphasised the fact that examples of evolutionary stasis are well known from the fossil record and that ideas of species undergoing rapid adaptations that were not specifically linked to environmental changes have been around since the 19th century.
Bokma argued that trait co-adaptation is an important source of evolutionary changes. He cleverly illustrated this argument by means of a Lego car; I always think that using something other than just slides in a presentation greatly improves the talk! He argued that changes in colour or shape attributes can happen “easily” over a short evolutionary period as an adaptation to environmental changes. For example, characteristics of fur in Vulpes vulpes/lagopusfoxes(the colour of the Lego car) or beak sizes in Darwin’s finches (the windscreen of the Lego car) have evolved to increase the fitness of the individuals living in particular environmental conditions.
But things get trickier when it comes to major changes or adaptations. Bokma illustrated this idea by trying to change the overall shape of the car (like going from a regular car to a truck). As soon as he started to remove one wheel, the car totally lost its fitness! For non-Lego fans he also gave the example of enzymatic reactions in endotherms: if just one enzyme evolves to be more active at a temperature different to normal body temperature, then there is no increase in fitness for the organism (and probably a decrease as that precise enzyme will be less efficient). It is only when all enzymes co-evolve to be more active at a different body temperature that there might be an increase of fitness for an individual. Bokma used this example to emphasise the irreversibility of evolution and how lineages can get stuck in “evolutionary dead-ends”. In response to H. Falconer’s comment to Darwin (1862) that the Elephant remains “unchangeable & unchanged”, Bokma argued that it might be “easy” (in an evolutionary way) to become an elephant but once you’re there, climbing into trees or crawling underground becomes tricky. So for these bigger evolutionary changes (the “jumps” in punctuated equilibrium theory), one needs a “genetic revolution”; evolving from a flightless theropod to a turkey involves co-adaptation of more than just locomotory characters. There is a strong irreversibility component to evolution.
While some people were kind of miffed by Bokma’s talk and started a polite but energetic debate involving genetic mumbo jumbo, I just thought that his talk was a great illustration of the problem of stasis and jumps in evolution. As Gene Hunt demonstrated, evolutionary stasis and jumps are clearly dominant in the history of life but they are trickier to explain than gradualist evolution. However, to be objective I have to agree with some of the general comments on Bokma’s talk; “So what? We already knew that.” It reminded me of S.J. Gould’s excellent review on part of this idea in the Panda’s Thumb (1980): The Return of the Hopeful Monster. So yes, we do already know about the ideas Bokma presented and he didn’t bring any staggering new insights but I believe he illustrated it with talent and brio! And perhaps I should mention that I can’t be completely objective here; I just love Lego, sorry.
Strange things are stirring from the deep. Creatures from myths and legends, the sea serpents of old, are descending upon our shores to warn us of impending doom. . .
Well, maybe not. But the news that not one but two mysterious oarfish have been found dead and beached in California recently has spread around the world, reminding people that the oceans still harbour creatures that are stranger and more alien to us than even the most hallucinogen-induced science fiction creations.
Oarfish are members of the genus Regalecus. There are three species, R. glesne, R. kinoi, and R. russelii, found in the Atlantic, Eastern-Central Pacific and Indo-Pacific respectively. They are the largest bony fish in the world and can reach over 11m in length. Very little is known about them. Unfortunately most of the published literature concerns their meristics which are important but make for incredibly dull reading.
Oarfish
Regalecus is one of two genera in the family family Regalecidae. The other genus, Agrostichyhtys, is monospecific and contains the species A. parkeri, commonly known as the ribbonfish or streamerfish. A. parkeri is even less well-known than the oarfish. Wikipedia says that it is known from only seven specimens, though I know this is an underestimate as I have personally seen and dissected two that are not included in that list and I’m sure there are more in museum and fisheries department basements around the world that just haven’t made it into the literature.
Agrostichthys parkeri
There are a number of reasons the Regalecids are so poorly known. They live in the open ocean and if caught at all it is usually as part of a commercial catch where scientific research is low down the list of priorities. Their delicate shape also means that they damage easily and often what is hauled on board is barely recognisable as fish, let along as a Regalecid.
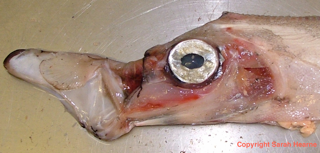
Another reason is that they elude scientific surveys as they are perfectly camouflaged and are almost impossible to see. I know this may sound crazy – an 11m long shiny silver animal with a gigantic eye surely can’t be hiding, it’d be like dressing up as a mirror-ball and expect to be able to blend into the background. But actually, that’s precisely what they are doing.
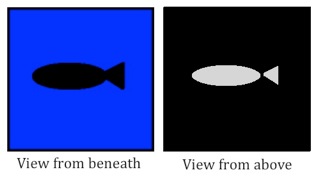
Their morphology has evolved for a very specific environment: the mesopelagic zone. This zone, colloquially termed the ‘twilight zone’, is the transitional part of the ocean, where light is still visible when you look up but there is nothing but blackness when you look below. A fish that is swimming in the water column will be seen as a black silhouette against the light when viewed from below or will stand out against the black of the deep when viewed from above.
This makes mesopelagic fish vulnerable to predation so they have evolved a number of countermeasures. The first is to be laterally compressed. This means that there is less of them to block or reflect any light. If you cannot compress laterally then a second option is to countershade – make your belly light and your black dark. Pelagic sharks, fish and even birds do this – it’s why penguins wear tuxedos.
Now that attacks from above and below have been dealt with, attacks from the side must next be tackled. For this, light contrast isn’t so much of a problem, but uniformity is as fish present a uniform block of colour against a dappled background. And this is where the silver comes in. The reflective surface of the fish, combined with their undulating body, help break up the light and allow it to merge into the background. If you don’t believe me watch this video of an oarfish in its natural habitat.
These adaptations mean they are able to successfully hide from all but the luckiest of predators. Yet life is more than just about avoiding being eaten. It’s also about finding food. This is incredibly difficult in the sparsely populated open oceans. and it is here that the giant eyes come in. Unlike the bathypelagic zone where eyesight is largely redundant, here in the mesopelagic zone there is just enough light to see by. You’ll notice the large pupil which has evolved to catch every last photon of light in order to detect potential prey, who, it’s worth pointing out, have evolved the same anti-predation adaptations as the Regalecids. If you watch the video you’ll see that oarfish sit vertically in the water column, looking up towards the ocean surface so they can fully exploit their excellent vision and spot anything passing overhead. Prey are few and far between, and are constantly on guard against being eaten. To catch anything stealth and speed are essential. And this is where their weirdest adaptation comes into play.
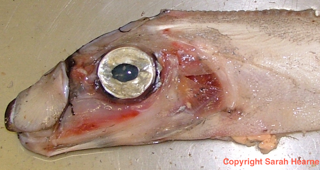
Before I continue, I want to point out that what follows is pure speculation as I can find nothing in the literature that discusses this, the most bizarre aspect of Regalecid morphology. I thought it was unique to A. parkeri but closer inspection of photos and videos (this is R. russelii) indicates this is a feature common to the family. I am talking about their mouth.
At rest there is little of note.
It’s a bit strange in appearance, almost beak-like, but fish have evolved all sorts of weird appearances. Nothing to worry about, maybe.
Think again.
The ‘beak’ extends! It forms a sort of funnel and we (my former colleagues and I) speculated that this might be to try and change the water pressure to help suck prey in. The narrow tube quickly opens up into the wider ‘pouch’ towards the back. This suggests there will be rapid changes in water pressure but without proper study, and live observations, this is all complete conjecture which is crying out for research.
As if all that isn’t enough to pique your interest, there are reports that the live fish give electric shocks:
“Jason McKenzie reported that when he picked up the fish . . . he experienced what he described as mild but quite distinct pulsed electric shocks that passed up through his hand into the forearm at intervals of about 10 s. A companion who independently held the fish reported the same sensation.”1
There is nothing coherent I can say about that; it just blows my mind!
It’s a real shame the media frenzy has been so superficial. There is so little known about these animals: basic biology such as how long they live, how often they reproduce, how they find mates, what they eat and what eats them is all unknown. These new specimens will undoubtedly be studied exhaustively. The media hype will have died down by the time the results are published, but I recommend keeping an eye out as they will add so much more to the understanding of these magnificent, elusive animals.
1 McDowall, R M & Stewart, A L (1999). Further specimens of Agrostichthys Parkeri (Teleostei: Regalecidae), with natural history notes. Proceedings of the 5th Indo-Pacific Fish Conference, Noumea, 1997. pp165-174. Séret, B & Sire, J-Y, eds. Sociéte Française d’Ichthyologie, 1999, Paris.
Author and Picture Credits: Sarah Hearne, hearnes[at]tcd.ie, @SarahVHearne
As I wrote in a previous post last winter, O’Leary et al. added their oar into the Placental Mammal origins debate. For anyone who missed that episode, they argued, with the backing of masses of morphological data, that placental mammal orders appeared right after the extinction of non-avian dinosaurs (also known as the explosive model). This was in opposition to two other views based on DNA data which argue that placentals appeared way before (long-fuse model) or slightly before (short-fuse model) the Mexican dinosaurs had to deal with some meteorite… Again, have a look at this previous post criticizing O’Leary et al.’s paper and how they “forgot” to use (ignored?) state-of-the-art phylogenetic inference methods.
While I was away feeding mosquitoes in Finland – and wondering whether the lack of fishes for dinner was due to my poor fishing skills or the absence of fishes in the river – Science published two new episodes of the placental saga. Of the two, Springer et al. took the decision to properly criticize the methods of O’Leary et al.’s work. Amongst their detailed methods review, they particularly underlined the inaccuracy of O’Leary et al.’s explosive model; such a hypothesis would imply that the early placental mammals had a rate of molecular change similar to that of retroviruses. For over ten years it has been widely accepted that molecular rates (i.e. the number of DNA changes that are transmitted to descendants) vary among lineages through time. Knowing that, one can estimate these rates (or call it speed if you’re more comfortable with that) of evolution by calibrating phylogenetic trees with fossils. So, in this case, the amount of evolution needed to evolve from the late Cretaceous (~65 myr) non-placental mammals to the first placental mammals (~58 myr) has to be as high as evolutionary rates more characteristic of retroviruses to realistically explain this evolution.
Herein lies the eternal debate between palaeontologists and molecular biologists. The former base their estimations on the morphological changes they can see in the fossil record (even though some, as O’Leary et al. also include molecular data) while the latter calculate their evolutionary rate estimations on the molecular changes that they infer from living species’ DNA. Fundamentally, each method is valid but they are describing slightly different things ; palaeontologists infer the rates of changes between morphospecies (i.e. species that are separated based on their morphology) while molecular biologists study the rates of changes between surviving genetic pools (i.e. the populations leading to living species). My guess is that the true evolutionary history (i.e. the morphological and molecular changes of all the populations –fossils and living– through time) is to be found somewhere between these two approaches.
And that’s what I think O’Leary et al. demonstrated in their response to Springer et al.’s comments. Through a kind of a dodgy answer in reply to the technical points that Springer et al. underlined as the “retrovirusesomorph” rates, O’Leary’s team reran the analysis and found that yes, maybe the explosive model is not very realistic regarding the molecular data but neither is the long-fuse model regarding the palaeontological data. So which one should we choose? Hmmm, why not just go for the middle way with the short-fuse model? OK let’s do that – without calling it a short-fuse model though (they called it an “explosive model” in figure 2-B but to my mind at least, it’s getting closer to the short-fuse one).
So all that for what? Nobody can either deny O’Leary et al.’s amazing work nor claim that the long-fuse model is realistic; the consensual short-fuse model remains pretty well supported among both moderate palaeontologists and molecular biologists. However, I still cherish this paper because it shows how I think good science should always work; find the two extreme scenarios and then study the median one…
This blog post was originally written and published here on the SoapBox Science blog. The Soapbox Science event showcases UK women in science to inspire the next generation of scientists by making science fun and accessible. This year it will take place on Friday 5th July 12-3pm on the Southbank in London. If you’re in the area why not go and take a look?
I’ve been touring international natural history museums as part of my PhD research. The “behind the scenes” aspect of each museum is fairly unchanging; row upon row of cabinets with some very unusual objects lurking within – the taxidermic tastes of some people just leave you wondering… Aside from the obvious dissimilarities in size, the major difference between the museums I’ve visited is in the style of their public exhibits. Like any other industry, museum exhibition styles are subject to fashion trends which reflect society’s interests and inclinations at the time of the exhibit’s creation. Visiting collections in Dublin, London, Washington DC, New York and Harvard University is an interesting trip through the evolution of natural history museums.
A visit to Dublin’s Natural History Museum should be treated as an historical trip back to the golden age of Victorian interests in the natural world. The museum’s first floor gallery is typical of the “classical” approach to natural history collections; collect and stuff as many animals as possible and cram them together in display cases to be admired. Scientific information is often limited to just species names and any niggling doubts about taxidermic accuracy should be ignored (have a look at the Orang-utan’s less than life-like features on your next visit). I love this old style, not only because you never know what odd creatures are lurking around the corner but also for its testimony to the collecting frenzies of 19th century naturalists.
A quick tour around London’s natural history museum gives a flavour of the evolution of exhibition styles; from display cabinets similar to Dublin’s “dead zoo” approach through to the ultra-modern and interactive Darwin Centre where you can watch a real-life scientist at work behind a glass screen (maybe not so far removed from a zoo after all?) The human biology exhibits lie somewhere in the middle of these two extremes. The interactive displays that allow you to re-live your time in the womb and explore how your senses work represent some of the earliest changes in exhibition planning; the move from passive presentation of natural objects to interactive attempts to inform and entertain museum visitors. These changing attitudes are documented in Richard Fortey’s excellent book about life at London’s museum.
In different ways, the American museums I visited also mix old and new approaches to museum exhibits. The Smithsonian Institute seems to have a taste for dynamic and life-like taxidermy. They have a giraffe drinking from a waterhole, a leopard hanging out with its kill in a tree and lions in the middle of taking down a water buffalo.
They’re the same species as can be found in Dublin or London but their action-style poses certainly adds a bit more realism to the exhibits. You just have to wonder about what weird contortions and odd framing apparatus must have been used to preserve these animals in mid-action poses for ever more.
The American Museum of Natural History has a different approach to adding a touch of life to their dead zoo. Many of their species are displayed in dioramas; recessed windows frame scenes from Savannah, tropical rainforest, desert and woodland life. The AMNH also has a cleverly designed and beautifully displayed wing where visitors can walk the vertebrate tree of life. Feeling somewhat Dorothy-esque, you follow the vertebrate phylogeny laid out on the ground, stopping en-route to see fossil or skeletal examples from all the major lineages. Instead of going down the “pull a lever/ push a button/ touch a screen” route of exhibition interactivity, the AMNH has enlivened the traditional natural history style by displaying their species in an evolutionary or ecological context which is far more interesting and informative for the visitor. They are certainly lovely scenes to admire, though I do admit that I spent most of my time wondering what happens when everything comes to life at night…
Although smaller in size, Harvard’s Museum of Comparative Zoology combines all the exhibition styles of its big city museum counterparts. There are dead zoo-style cases of stuffed animals, ecological dioramas of species found in New England forests and high tech, interactive touch screens to explore the tree of life (a lovely tool but not completely up to date – because I’m pedantic I had to check how they classified tenrecs…) The added benefit of the MCZ is their inclusion of exhibits based on current and previous research of the museum and affiliated staff. They have specialised, themed exhibits based on their research expertise, I particularly loved the displays about the evolution of animal colouration and camouflage based on research from the Losos lab. Despite its limited size, the MCZ combines all major aspects of the evolution of museum exhibits; from static stuffed animals through to interactive attempts to inform visitors and clear links with active, current scientific research.
There’s clearly a huge variety in natural history museum exhibits and it’s interesting to see how each institution tackles the task of preserving their tradition while still continuing to keep a pace with the demand for new and more exciting exhibits. Our own TCD Zoology museum is no exception to this evolving museum trend as we prepare to welcome our first public visitors of the summer in just a few weeks’ time. I look forward to sharing some our own quirky museum stories; just how did our elephant, Prince Tom, fit in through the door?
Imagine you’re stuck in the desert, your plane has crashed and you’re trying hard to fix it. Then a child pops up out of the blue and asks you straight out “If you please – draw me a dino…”. Now let’s say you do as Antoine de St-Exupéry and take up the challenge without asking too many questions. How would you draw that dino? I guess it depends on when you were asked the question.
Let’s go back through the history of drawing dinosaurs. The pictures I grew up with were the ones from Jurassic Park who came directly from the last dino-revolution started by Ostrom and Bakker‘s work (especially with the publication of Dinosaures Heresies, in 1986). This vision was then heavily popularized by the three (nearly four?) blockbuster movies we all know and love…
Since the late 1990’s, the increased availability of formerly highly expensive techniques such as CT scans or synchrotrons, has helped to understand dinosaurs better than ever. Led by new discoveries from vast, previously unexplored deposits, some of the most interesting work from recent times shows an even closer link between birds and dinos than we previously thought (see last week’s Science NOW). So I believe (and hope) that the next generation will grow up with the pictures of dino-chickens and see their lunch time chicken wings as true theropod meat…
It is always really interesting to look at all the work that has been done and presented to the public through dinosaur pictorial art; from the weird/funny starts in the first half of the 19th century to the modern, highly accurate representations of today (the French artist Alain Bénéteau is just one example among many). As a nice example, have a look at the pictorial evolution of the second oldest scientifically described dinosaur: the Iguanodon.
But here I’d like to emphasize my love for what I think was the “golden age” of dinosaur pictorial art. I obviously want to refer to the work of artists like Charles R. Knight (1874:1953), Zdeněk Burian (1905:1981 – have a look at this awesome online gallery) or Rudolph F. Zallinger (1919:1995). Their beautiful and (for their time) highly accurate scientific artwork was crucially import for bringing palaeontology into the public eye. This “golden age” was made possible by the upgrading of palaeontology to the status of a true science and the general acceptance of Darwin’s theory. Public interest in palaeontology at this time was also fueled by new fossil discoveries from expanding European colonies and the American frontier eventually leading to the most epic palaeo-story ever: the Bone Wars (soon to be seen on HBO)!
I refer to this period (second half of the 19th century and first half of the 20th) as a “golden age” but that does not mean that it was the peak of palaeontological discovery or interest. Our palaeo-knowledge has never been richer. Dinosaurs are no longer merely unknown beasts from an ageless past. They are now placed in accurate phylogenetic frameworks and are just one of the many extinct tetrapod groups which we can now link to extant biodiversity. However, identifying birds as living remains of the dinosaur lineage does diminish dinosaurs’ mightiness. They are no longer the perfect romantic group of fossils: giant monsters that ruled the earth for over 150 million years before being completely wiped out by a single meteorite that cleared the way for us to evolve and exist. Even if I actively try to fight against this simplistic view of the History of Life, I have to admit that it is the one that brought me into palaeontology, not the chickens I used to keep in my parents’ garden… So, although I have no drawing talent whatsoever, because I think that dinosaurs are still awesome but lacking the mightiness they deserve, I’ll draw that little prince something like this:
A sketch from De la Beche (1930) titled: Dr M[antell] in extasies at the approach of is pet SaurianThis post was inspired by the excellent “Dinomania” chapter of Gould’s Bully for Brontosaurus 1991, by a master’s project done with F.Barbiere, S.Enault and B.Ramassamy and by the excellent blogs which can be found about this subject such as here, here or here.
My former PhD student, Luke McNally and I authored a paper published recently showing how “Cooperation creates selection for tactical deception”. Using a combination of mathematical models and analysis of empirical data from 24 primate species, we show that acts of deception are more likely to occur when the individuals in the group show greater cooperation. In other words, deception and cooperation go hand-in-hand. Perhaps not a surprising result, as Rob Brooks recently pointed out in a very accurate and nice blog post on our paper, but the evolutionary forces that might maintain deception in society have not been previously described.
We have enjoyed some media coverage with this paper, including some international science slots, a bit of national radio and Rob’s blog post. I take some mixed pleasure in the fact that a creationist website picked up on both our paper and Rob’s post. Its something of a tongue-in-cheek achievement to have caught their eye given my total opposition to creationism in all its forms. I’m also quite proud to have earned a “Darwin baloney” award (which I might add to my website as a badge of honour assuming I’m not infringing copyright). Im also intrigued to have the mental disorder “Yoda Complex” bestowed upon me by this group, even if it is not the Urban Dictionary definition but rather their own invention because “because we thought of it independently” (Editor’s comment in http://crev.info/2013/05/evolutionists-confess-to-lying/). So happy with this flattery than I now tweet under @yodacomplex.
Ordinarily I would steer clear of getting sucked into arguing with such groups, but their article just annoys me. I’m even more annoyed that I can’t reply to their post on their site without signing into their site, and registering with them is a bridge too far. Equally frustrating is their anonymity which makes directing my counter-arguments somewhat indirect.
The consequence of their argument is that “if lying evolved… how are readers to know who is telling the truth?” which leads them to the title “evolutionists confess to lying” (http://crev.info/2013/05/evolutionists-confess-to-lying/).
The basis of their argument goes:
“Imagine a liar so skilled, he convinces his listeners that he is 100% against the worst dishonesties in politics, public relations and propaganda. He tells you he wants to achieve enormous social good to provide a better understanding of how lying evolves. Now, add to it that he is self-deceived. Doesn’t his credibility implode? How could one possibly believe a word he says?”
How can one believe what a person says? This is exactly why we have science. Our results are open for all to examine and check. The results might be incorrect (but we are confident in our analyses), but until someone shows us exactly where we have gone wrong, then we can take them as being a true and fair reflection of our study system. Our mathematical model shows under what circumstances deception (lying) can be sustained in an evolutionary sense in any society subject to a cooperative based reward system (in this case a system governed by the Iterated Prisoner’s Dilemma). The prediction from the model is that mechanisms that might enforce cooperation (such as only cooperating with other co-operators and spurning those who cheat) create a niche where lying can profit and proliferate. Our analysis of data from 24 primate species backs up our theoretical model, showing that the more likely a species is to engage in cooperative acts, the more likely deception is to occur in their society.
The creationist author goes on to make a major error in interpreting the whole basis of the study of the evolution of social behaviour.
“In the evolutionary world, there is no essential difference between cooperation and deception. It’s only a matter of which side is in the majority at the moment.”
This is just plain incorrect, and is the entire basis for their spurious argument. In the study of social behaviour (irrespective of evolution) there is indeed a fundamental difference between cooperation and deception (although I think they really mean defection here, with deception being a means to hide ones defection in the wording of the Iterated Prisoner’s Dilemma). In the Iterated Prisoner’s Dilemma, and related games like the Snowdrift Game, cooperation is the act of assisting another individual so as to share a reward. Defection on the other hand is the act of cheating on the other person in the game so as to walk away with the entire reward for themselves. It is absolutely not a “majority” based definition. Deceivers in our model try to trick co-operators so as to walk away with their share too by convincing them that they intend to cooperate. The kool-aid scenario that follows in that blog post is just not relevant since it invokes a semantic argument about how the players choose to define cooperation and defection that is simply not present in these evolutionary models of social behaviour. All the author has done is to flip the labels of co-operators and defectors. The outcome of their scenario would be that the poisoners (who are actually the defectors as per any sensible definition of their behaviour in cooperative game) would kill all the co-operators leaving only themselves. Indeed, this matches the fundamental prediction of the evolutionary models which offer “defect all the time” as a consistent stable end-game scenario. It is the goal of most evolutionary studies of social behaviour to learn what mechanisms exist in societies that mean we don’t get stuck here, since it is clear that many primates, including humans, have a much more cooperative society than that depressing outlook.
“Since all these evolutionists believe that lying evolved as a fitness strategy, and since they are unable to distinguish between truth and lies, they essentially confess to lying themselves. Their readers are therefore justified in considering them deceivers, and dismissing everything they say, including the notion that lying evolved.”
This is the rather annoying consequence of their incorrect logical arguments. We can and do distinguish very clearly in our models and reasoning between truth and lies – at least in these models we do. Also, just because we point out that lying can have an evolutionary selective advantage (which is hardly surprising), surely doesn’t make us liars? I can’t see what the mechanism there could possibly be.
Just to end, I have to say that it is really difficult not to ridicule this type of article. The reasoning is just so off-the-wall, based on a manipulation of what science is all about, and with a really nefarious motivation running through it of debunking science for the true believers. I did laugh, I did sneer, (and I did take @yodacomplex as a twitter account, and I love it); but, I have tried here to avoid sneering since they use that against us (see the comments under their article). In fairness though, giving us a “Darwin Baloney” logo, and administering a mental health disorder on us (even if they made it up themselves) is pretty much name calling and sneering in my book – even if I am rather flattered to have acquired their attention.
Nature News published a new post about our origins. It’s promoting Stevens et al’s 2013 paper that published the description from two new granddads/grandmas in our already complicated family tree. These guys, Nsungwepithecus and Rukwapithecus (it’s not that hard to pronounce, try it) are considered by Stevens and his team as the oldest crown Catarrhines – [Google translate palaeo-primatish to English: “as closely related as the ancestors of you and your cousin the proboscis monkey (we all knew there were some facial similarities!)”]. Technically speaking, these fossil discoveries pushed the origin of modern Catarrhines back from 20 to 25 million years ago, a date which is closer to molecular results (25-30 mya). Astonishing eh? Well it is for me but there’s another reason why I wanted to talk about this paper: gaps.
As many people might know, the fossil record contains some serious lacunas; thick layers of rock containing either very few or no fossils at all. One example mentioned in this paper is the scarcity of fossils from the Oligocene period; very few bones for palaeontologists to gnaw… The Oligocene was, however, a period of massive changes, in climate and all that stuff but also the time when placental mammals evolved from their primitive to modern forms. So the lack of fossils from this time is always frustrating when you want to understand macro-evolutionary patterns. But why have we found so few Oligocene fossils until now? Of the many explanations Stevens et al. suggest that “Possible reasons […] include limited deposits of appropriate age, particularly […] below the equator, complicated by densely vegetated topography in more tropical environments”. And that’s why I find this article so exciting! Traditionally, fossil primates were excavated in deserts or northern latitudes, which is typically where they are fairly rare nowadays! Therefore, I’m happy to see that there is a true effort being made to look for fossils in biodiversity hotspots such as Tarzan’s neighborhood (or the Tanzanian Rukwa Rift Valley in this paper) where these new primates were discovered.
I think one of the most important things to come from this paper is that it’s proof of a real effort to look for the fossils in the true biodiversity hotspots which, I’m sure will lead to far better comprehension of modern mammalian history. These new primates came from Africa but people are also working in Peruvian jungle and interesting new discoveries are not limited to just primates…
The Toco Toucan (Ramphastos toco) is the largest species in the toucan family but not only that, it has the largest bill relative to body size of all birds. As with most things in Zoology the function of the bill has been hotly debated, even Darwin himself weighed in with an explanation of his own. He thought that the exaggerated size of the bill may have been due to sexual selection. Seems a little extreme though, especially when you consider all the adaptations for flight birds already exhibit (see here for the basics). Why then would natural selection begin to select individuals with heavy large beaks, surely the extra matings acquired due to the size of your “birdhood” would be offset by your reduced capacity for flight. Right?
Well that depends, what if there were other benefits to having this huge bill. Like for example thermoregulation. Like other enlarged body parts used for thermoregulation, like for example the enlarged ears of many desert dwelling mammals, the bill of the toucan is highly vascularised (supplied with blood vessels) and it seems the toucan has the ability to control the amount of blood flowing to the blood vessels around the bill.
Thermoregulation is somewhat of a hot topic (if you’ll pardon the pun), recently there has been some suggestion that the plates and spikes of the Stegosaurus may have been candidates for thermoregulatory function, where they had been previously thought to have been for defence. Similarly and somewhat more bizarrelythe long neck of many animals both extant and extinct has been discussed as a possible means of thermoregulaltion in this wonderful article by Wilkinson and Ruxton (2011).
So the long and the short of it? As with many aspects of the animal kingdom, without wanting to blunt Occam’s razor the simplest answer may not always be correct.

























{kind=link}