Nature News published a new post about our origins. It’s promoting Stevens et al’s 2013 paper that published the description from two new granddads/grandmas in our already complicated family tree. These guys, Nsungwepithecus and Rukwapithecus (it’s not that hard to pronounce, try it) are considered by Stevens and his team as the oldest crown Catarrhines – [Google translate palaeo-primatish to English: “as closely related as the ancestors of you and your cousin the proboscis monkey (we all knew there were some facial similarities!)”]. Technically speaking, these fossil discoveries pushed the origin of modern Catarrhines back from 20 to 25 million years ago, a date which is closer to molecular results (25-30 mya). Astonishing eh? Well it is for me but there’s another reason why I wanted to talk about this paper: gaps.
As many people might know, the fossil record contains some serious lacunas; thick layers of rock containing either very few or no fossils at all. One example mentioned in this paper is the scarcity of fossils from the Oligocene period; very few bones for palaeontologists to gnaw… The Oligocene was, however, a period of massive changes, in climate and all that stuff but also the time when placental mammals evolved from their primitive to modern forms. So the lack of fossils from this time is always frustrating when you want to understand macro-evolutionary patterns. But why have we found so few Oligocene fossils until now? Of the many explanations Stevens et al. suggest that “Possible reasons […] include limited deposits of appropriate age, particularly […] below the equator, complicated by densely vegetated topography in more tropical environments”. And that’s why I find this article so exciting! Traditionally, fossil primates were excavated in deserts or northern latitudes, which is typically where they are fairly rare nowadays! Therefore, I’m happy to see that there is a true effort being made to look for fossils in biodiversity hotspots such as Tarzan’s neighborhood (or the Tanzanian Rukwa Rift Valley in this paper) where these new primates were discovered.
I think one of the most important things to come from this paper is that it’s proof of a real effort to look for the fossils in the true biodiversity hotspots which, I’m sure will lead to far better comprehension of modern mammalian history. These new primates came from Africa but people are also working in Peruvian jungle and interesting new discoveries are not limited to just primates…
I’d like to ask the question many paleontologists have to face when they (foolishly) venture out of their museum storage: “So you’re studying fossils right? But what will that bring to the people? A cure for AIDS?”. There are many possible answers from a punch in the face to more mature responses. But I was recently asking myself the question from a biologist’s point of view: “What can biologists really do with the fossil record?”. Well obviously, we can use it to recreate and understand the history of our planet (like in Nature last week) or to do use some nice methods in trying to understand ancient ecosystems. People even might feel lyrical and do some serious work on paleo-poetry! But all of these guys are paleontologists right? They live in their museums and only go out for a movie once every 10 years… How about the other biologists?
Think about it, when ever you’re studying any organism, it is obvious (thanks to this bearded ape) that they had a 3.5 billion year history behind them. Ignoring that might lead to a misunderstanding? As an example, I’d like to use my favorite PhD-presentation example: the crocodiles. When we talk about crocodiles, we automatically think about the few species of big lizard that live in rivers in the tropical/sub-tropical latitudes. But, after a quick look at the history of our planet, the only description that is more or less correct is “lizard” (archosaurians to be more precise). Crocodiles are composed of many species (8 genera today – soon to be 6 – but >70 in prehistoric times) that lived in rivers as much as in the sea, on the ground or even sometimes in trees and in tropical to temperate climates (remains of crocs were found in Normandy – France).
Well maybe that’s just because of this group. But if you think about it, many other groups have ecological or evolutionary features that becomes truly astonishing once you take into account their full history. For my PhD I decided, with Natalie, to look at this fun fact (life existed before yesterday and the people studying it don’t always focus on dinosaurs) through primates. My idea is to combine extant data based on DNA with extinct data based on morphology to have an integrative tree of all primate history. I agree that this sounds a bit too easy and naive, (the method is a bit more complex) and I’ll probably end up with something more humble. However I think the primates can be a good example to illustrate the point about the hidden diversity among extinct groups. The primate fossils are not dramatically different than the extant once (unlike crocs, there were no pelagic primates) but they still show some really interesting features, for the macroecology side, combined extant and extinct primates show massive variation in body mass in some groups (lemurs) but very few variations in others (tarsiers). Or on the macroevolution side, such an integrative tree could provide some further understanding to the old debate of primate origins! Well at least I hope so. For now I’m just comfortable with eating some burgers with a diet coke and a gun in a pickup truck while I’m scanning some primates in the Smithsonian Institution in Washington DC.
*by parasites here I am referring to all kinds of infectious disease causing agents including bacteria, viruses, fungi, protozoa, helminths and arthropods.
Why do we care about primate parasites?
Many of the most devastating infectious diseases in humans have origins in wildlife. For example, the global AIDS pandemic originated through human contact with wild African primates and influenza viruses circulate among wild bird populations. These are not only historical occurrences. Recently, for example, rodents were identified as the source of a Hantavirus outbreak in Yosemite National Park, USA . As human populations continue to expand into new areas and global changes in temperature and habitat alter the distributions of wild animals, humans around the world will have greater contact with wildlife. Thus, understanding which infectious agents have the potential to spread from animals to humans is crucial for preventing future human disease outbreaks.
Many efforts are being made to collate information on wildlife and human diseases. Much of my research (which I will blog about when I get chance!) uses an amazing database known as the Global Mammal Parasite Database or GMPD for short. Every time a paper is published which contains details of parasites found in either primates, carnivores or ungulates, the information is added to the database. As much data as possible is recorded, including the species infected, the type of parasite, the prevalence of the parasite, and the geographic location of the study. Prof. Charles Nunn and his colleagues have been collecting data for the GMPD since around 2005 and it currently contains around 6000 records for primates alone. This definitely makes it the most comprehensive dataset of primate parasites in existence.
The GMPD sounds amazing…so what’s the problem?
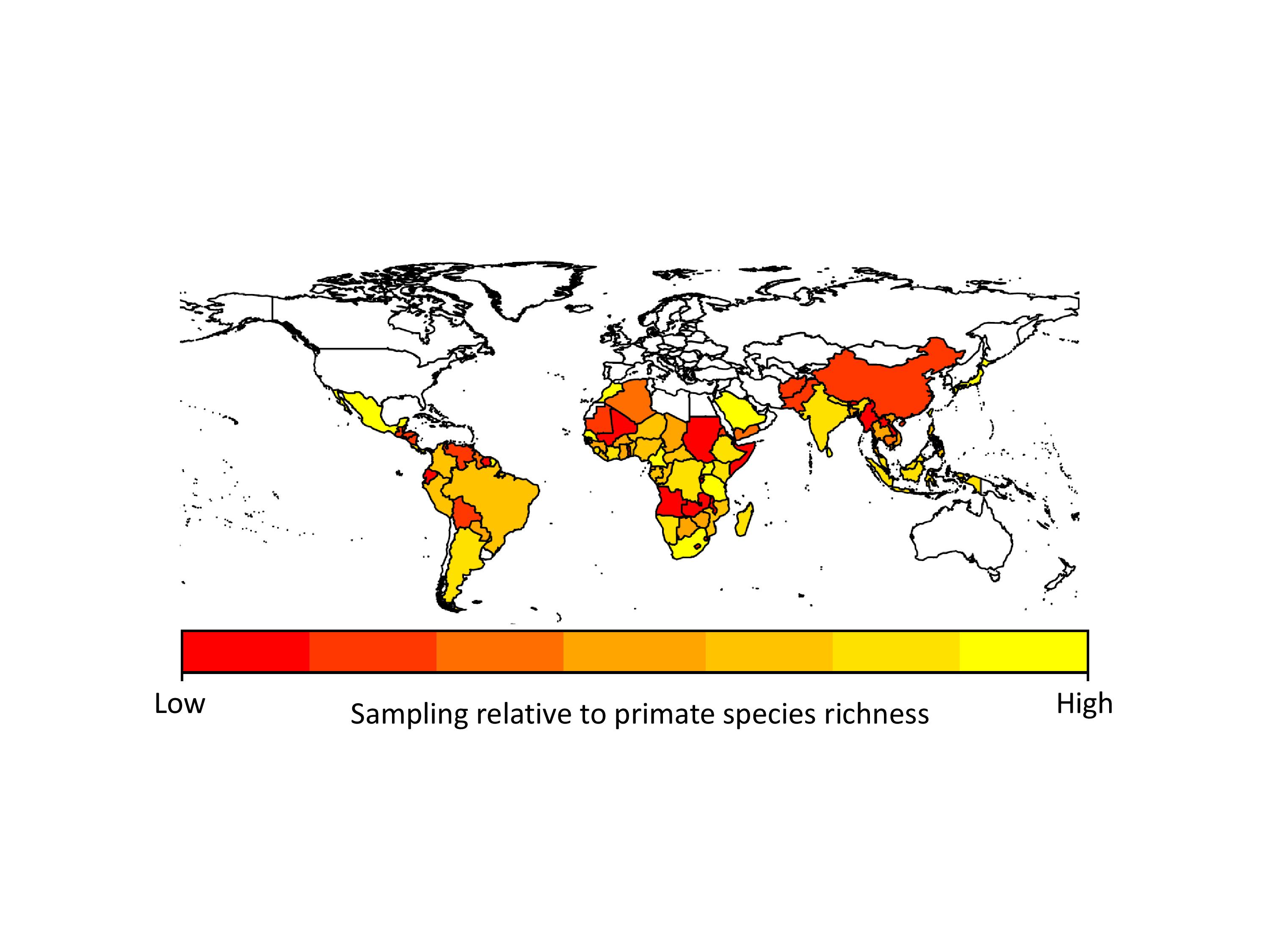
The problem with the GMPD (and this is a feature of virtually all datasets) is that there is sampling bias. Certain primates are sampled for parasites much more frequently than others. Chimpanzees, for example, are sampled for parasites all the time, whereas species such as tarsiers are sampled much less often. This has the effect of making it look like chimpanzees have far more parasites than tarsiers, simply because they have been sampled more often. In analyses using the database we usually deal with this problem by adding sampling effort into our models, so we give less emphasis to high numbers of parasites in primates we have lots of samples for. Unfortunately this problem is also evident when we look at parasites (things like malarial parasites are often sampled because of their importance to human health) and geographic regions (areas with primate research stations are sampled far more regularly than more remote regions). If we hope to use the GMPD data to make reliable predictions about future risks to humans, we need to identify gaps in our knowledge of primate parasites.
So what did you do?
Without going into the technical details, we looked across the primate phylogeny and primate geographic ranges to identify gaps in our knowledge, and used statistical models to investigate what factors led to primates and geographic areas being relatively well- or relatively poorly-sampled. We also used species accumulation curves to extrapolate parasite species richness for primates.
Where are the gaps in our knowledge?
We found that apes (chimpanzees, gorillas and orangutans) were generally better-sampled than other primates, but there was incredible variation in sampling among all other major primate groups. Apart from apes, the primates that researchers appear to sample most are the species they encounter most often, i.e., widespread, terrestrial, diurnal species. However, some primates were sampled more often because they are already intensively studied for other research, because they live in frequently visited field sites, or because of their importance in medical research. Across countries, we found that in general, parasite sampling is highest in countries with more primates to sample. We expected that the GDP of the countries would also affect sampling effort, with wealthier countries having more money for disease monitoring. However, we found no evidence for this in our analyses, probably because most research on primate diseases is not funded by the country in which the research takes place.
Sampling effort for primate parasites across the world. Poorly-sampled countries are in red, and better sampled countries are in yellow.
When we extrapolated parasite species richness values we found that even within our best-sampled primates and countries, we are missing a lot of parasites. On average we predicted that 38-79% more parasite species than currently reported in the GMPD should be found in our best sampled primate species, and 29-40% more parasite species than currently reported in the GMPD should be found in our best sampled countries. This emphasizes exactly how poor our sampling is across all primates and countries. Another concern is that although viruses make up only 12% of the parasites in our dataset, viruses arguably present the greatest zoonotic disease threat to humans because their fast rates of evolution should allow them to easily adapt to new hosts.
What next?
Identifying parasite sampling gaps across primate species and geographic regions is only the first step; we need to find strategies to minimize these sampling gaps if we are to predict which primate diseases may emerge in humans. One solution is to set research priorities based on the sampling gaps, for example, by focusing effort and funding on relatively poorly-sampled primate species, arboreal primates, those with small geographic ranges, or those found in relatively poorly-sampled regions of South East Asia, Central and Western Africa, and South America.
Focusing on relatively poorly-sampled primate species and areas may improve our general understanding of primate parasites, but it is only one factor in predicting risk to humans. For example, hosts are more likely to share parasites with their close relatives than with more distant relatives. Thus, continuing to focus our sampling efforts on parasites of our closest relatives (chimpanzees, gorillas and orangutans) may provide the greatest return in the case of risks to humans. This is particularly important because we found that chimpanzees are expected to have 33-50% more parasites than currently found in the GMPD. In addition, ecological similarities also influence parasite sharing among primates, and humans share more parasites with terrestrial than arboreal primate species. As with sampling effort, this probably reflects higher contact rates among humans and terrestrial primates compared to arboreal primates. As a related issue, a host living at higher density is expected to have higher prevalence of parasites and may have more contact with human populations or our domesticated animals, thus increasing opportunities for host shifts to humans. The large numbers of zoonotic emerging infectious diseases with rodent or domesticated animal sources also highlight the importance of rates of contact and host density for disease emergence in humans.
In conclusion Sampling effort for primate parasites is uneven and low. The sobering message is that we know little about even the best studied primates, and even less regarding the spatial and temporal distribution of parasitism within species. Much more sampling is needed if we hope to predict or prevent future emerging infectious diseases outbreaks.
I spoke before of how to use mathematics to convey an idea in biology. Here, I’ll take a different tack and discuss a paper in which the author makes his argument with naked English. The author in question is Nicholas Humphrey who in his famous paper ‘The social function of the intellect’ draws a wonderful metaphor of Mother Nature as an economist,
“It is not her habit to tolerate needless extravagance in the animals on her production lines: superfluous capacity is trimmed back, new capacity added only as and when it is needed”.
His metaphor serves as an introduction to the puzzle of the seemingly unnecessarily inflated intellects of some animals, notably humans.
Humphrey questions if such a highly developed intellect is really necessary for invention. The ability to produce tools is generally not a result of deductive reasoning or creative thought but rather follows from aping other individuals or pure trial and error learning. The intellect must have some other function in his estimation and in the end, he proposes that it is as a social glue. The complex interactions that arise out of the social milieu require some serious intellectual horsepower,
“[S]ocial primates are required by the very nature of the system they create and maintain to be calculating beings; they must be able to calculate the consequences of their own behaviour, to calculate the likely behaviour of others, to calculate the balance of advantage and loss – and all this in a context where the evidence on which their calculations are based is ephemeral, ambiguous and liable to change, not least as a consequence of their own actions.”
Calculating the consequences of your own behaviour is one thing but understanding that others around you have motivations of their own is a huge leap in understanding. All of this is done without ever having direct access to the subjective thoughts, motives, and desires of another person. Understanding the reasons for understanding is even more impressive and Humphrey’s paper has rightly influenced the theories of scientists since its publication. Most recently a study in the school that mechanistically linked sociality and selection for intelligence.